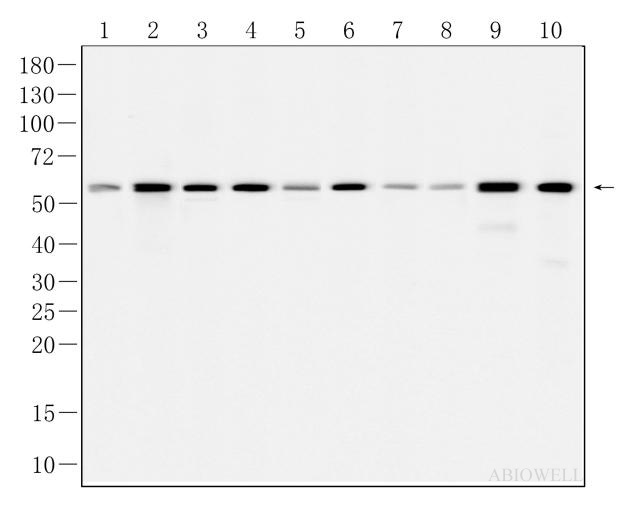
Ki67 Recombinant Rabbit Monoclonal Antibody





-
-
- 20μL
- ¥620
- 有库存
-
- 50μL
- ¥1250
- 有库存
-
- 100μL
- ¥2200
- 有库存
Product Details
| Host Species: Rabbit | Reactivity: Human,Mouse,Rat | Molecular Wt: 345-395 kDa | |
Clonality: Monoclonal | Isotype: IgG | Concentration: 1 mg/ml | ||
Other Names: MKI67; Antigen KI-67;Ki-67 | ||||
Formulation: Liquid in PBS containing 50% glycerol, 0.5% BSA and 0.02% sodium azide. | ||||
Purification: Affinity-chromatography | ||||
Storage: -20°C,1 year | ||||
Applications
| IHC-P: 1:5,000-1:15,000 IF: 1:500 | |||
Immunogen Information | Gene Name: MKI67 | Protein Name: Antigen KI-67 | ||
Gene ID:
| SwissPro:
| |||
Immunogen: Synthetic peptide within human Ki67 aa 1,040-1,080. | ||||
Specificity: Ki67 Monoclonal Antibody detects endogenous levels of Ki67 protein. | ||||
| Product images | |
|
|
Fig : Immunohistochemical analysis of paraffin-embedded rat-spleen tissue with Rabbit anti-Ki67 (AWA10320) at 1/100 dilution. The section was pre-treated using heat mediated antigen retrieval with Sodium citrate buffer (pH 6.0) for 20 minutes. The tissues were blocked in 3% H2O2 for 15 minutes at room temperature, washed with ddH2O and PBS, and then probed with the primary antibody (AWA10320) at 1/100 dilution for 1 hour at room temperature. The detection was performed using an HRP conjugated compact polymer system(ABIOWELL, AWI0629). DAB was used as the chromogen. Tissues were counterstained with hematoxylin and mounted with DPX. |
|
|
Fig : Immunohistochemical analysis of paraffin-embedded rat-testicle tissue with Rabbit anti-Ki67 (AWA10320) at 1/100 dilution. The section was pre-treated using heat mediated antigen retrieval with Sodium citrate buffer (pH 6.0) for 20 minutes. The tissues were blocked in 3% H2O2 for 15 minutes at room temperature, washed with ddH2O and PBS, and then probed with the primary antibody (AWA10320) at 1/100 dilution for 1 hour at room temperature. The detection was performed using an HRP conjugated compact polymer system(ABIOWELL, AWI0629). DAB was used as the chromogen. Tissues were counterstained with hematoxylin and mounted with DPX. |
|
|
Fig : Immunohistochemical analysis of paraffin-embedded Mouse-large intestine tissue with Rabbit anti-Ki67 antibody (AWA10320) at 1/200 dilution. The section was pre-treated using heat mediated antigen retrieval with Sodium citrate buffer (pH 6.0) for 20 minutes. The tissues were blocked in 3% H2O2 for 15 minutes at room temperature, washed with ddH2O and PBS, and then probed with the primary antibody (AWA10320) at 1/200 dilution for 1 hour at room temperature. The detection was performed using an HRP conjugated compact polymer system(ABIOWELL, AWI0629). DAB was used as the chromogen. Tissues were counterstained with hematoxylin and mounted with DPX. |
|
|
Fig : Immunohistochemical analysis of paraffin-embedded Mouse-heart tissue with Rabbit anti-KI67 (AWA10320) at 1/800 dilution. The section was pre-treated using heat mediated antigen retrieval with Sodium citrate buffer (pH 6.0) for 20 minutes. The tissues were blocked in 3% H2O2 for 15 minutes at room temperature, washed with ddH2O and PBS, and then probed with the primary antibody (AWA10320) at 1/800 dilution for 1 hour at room temperature. The detection was performed using an HRP conjugated compact polymer system(ABIOWELL, AWI0629). DAB was used as the chromogen. Tissues were counterstained with hematoxylin and mounted with DPX. |
|
|
Fig : Immunohistochemical analysis of paraffin-embedded Rat-heart tissue with Rabbit anti-KI67 (AWA10320) at 1/800 dilution. The section was pre-treated using heat mediated antigen retrieval with Sodium citrate buffer (pH 6.0) for 20 minutes. The tissues were blocked in 3% H2O2 for 15 minutes at room temperature, washed with ddH2O and PBS, and then probed with the primary antibody (AWA10320) at 1/800 dilution for 1 hour at room temperature. The detection was performed using an HRP conjugated compact polymer system(ABIOWELL, AWI0629). DAB was used as the chromogen. Tissues were counterstained with hematoxylin and mounted with DPX. |
-
-
- 20μL
- ¥620
- 1-3个工作日
-
- 50μL
- ¥1250
- 1-3个工作日
-
- 100μL
- ¥2200
- 1-3个工作日
-
相关产品
-
Cytokeratin 15 Recombinant Rabbit Monoclonal Antibody
HMGB1 Recombinant Rabbit Monoclonal Antibody
SQSTM1/p62 Mouse Monoclonal Antibody
Vimentin Mouse Monoclonal Antibody
xCT/SLC7A11 Recombinant Rabbit Monoclonal Antibody
SQSTM1/p62 Recombinant Rabbit Monoclonal Antibody
Caspase-8 Recombinant Rabbit Monoclonal Antibody
Ki67 Recombinant Rabbit Monoclonal Antibody